|
Dendropsophus ebraccatus Cope, 1874
 |
|
|
Family: Hylidae
Hourglass Treefrog, more...Hourglass Treefrog, Painted Treefrog, Painted Treefrog (es: Rana-de Árbol Amarillenta, Rana-de Árbol Amarillenta)
[Hyla ebraccata Cope, 1874]  Justin Touchon |
Adult: Species description based on Duellman (1970), Duellman (2001) and Savage (2002). Small, yellow treefrogs. Dendropsophus ebraccatus exhibits sexual dimorphism. Males are smaller than females: males to 28 mm; females to 37 mm. Snout-vent length, head width and tympanic membranes are all larger in females but laryngeal components are larger in males (McClelland et. al. 1993). An extensive description of the adults and tadpoles can be found in Duellman (2001). Dorsal: Dorsal coloration changes from vibrant yellow at night to pale yellow or cream during the day. Many specimens have a brown stripe running laterally from the tip of the snout through the eyes to the rear of the head. Dorsal surface with variable dark brown markings. Duellman (1970) recognized four dorsal patterns: plain, spotted, triangle, or hour-glass. Intermediate patterns have not been observed. The dorsal skin is smooth. Ventral: Creamy white. Concealed surfaces: Thighs are uniformly yellow with no bright coloration in the concealed portions. Eye: Iris brown to reddish bronze, sometimes tan or yellow. Pupil horizontal. Extremities: Foot moderately to extensively webbed. Toes with large terminal discs. Breeding season: The breeding season is largely concordant with the rainy season. In Panama, the breeding season lasts roughly from May to November (Fouquette 1960) or even January (Duellman 1970), depending on the duration of the rainy season. Large breeding choruses form at the edges of marshes and ponds. For females, time between reproductive bouts may be a little as 10 days (Wells 2007). Egg: Dendropsophus ebraccatus lay relatively large clutches of small eggs (Wells 2007); however the number of eggs per clutch is quite variable (Duellman 1970, Roberts 1994, Savage 2002). Eggs are laid terrestrially on leaves overhanging water (Duellman 1970). In Gamboa, Panama,eggs may are laid aquatically onto submerged or floating vegetation (Touchon and Warkentin 2009). Mutiple clutches of eggs are laid by a single female in a night (Touchon and Warkentin 2008). Undisturbed eggs hatch in approximately 3 ½ days. Tadpole: Tadpoles develop in ponds where they can often be dipnetted from aquatic vegetation or leaf litter on the bottom (Touchon pers. comm.). Tadpole bodies are mostly dark brown flecked with gold above, with a prominent black eye bar. The ventral surface is white. Tadpoles have large pigmented areas on the tail and develop bright red coloration in the presence of aquatic invertebrate predators (Touchon and Warkentin 2008a). Unlike species that are adapted to more ephemeral larval habitats, D. ebraccatus tadpoles cannot survive for very long out of water (Valerio 1971). Metamorph juvenile: Metamorphs emerge in as little as 6 weeks but duration of the larval stage varies considerably depending on pond temperatures (Touchon pers. comm.). Time to maturity and lifespan in the wild are unknown. Habitat: Humid, lowland forest to 1600 m. This species may also occur in areas that have been cut extensively (Duellman 1970). Adults most likely live in the forest canopy, although some individuals have been found in leaf litter in the buttresses of trees (Whitfield and Pierce 2005). Individuals have also been found in bromeliads (Duellman 1970). During the breeding season, individuals may be found on vegetation near ponds, marshes, and temporarilly flooded areas at night. Call: A prolonged "wreek" or "creek" followed by zero to several "eeks" (Guyer and Donelly 2005, Ibanez et al 1999). Some species of frogs emit rain calls in the daytime during or before rain showers. The rain call of D. ebraccatus sounds like the short primary note of the mating call (Duellman 1970). Karyotype: 2N = 30 (Duellman and Cole 1965, Kaiser et al 1996). Evolotion: Ohmer et al (2009) found divergence in body size, colour pattern frequencies, and male advertisement call among genetically distinct populations of D. ebraccatus in Panama and Costa Rica. Patterns of genetic and phenotypic variation differ from those of Agalychnis callidryas (Robertson 2008, Robertson et al 2009). Type locality: "region of Nicaragua" (Dunn and Stuart 1951) Physiology: The vocal sac of D. ebraccatus is not a cavity resonator (Rand and Dudley 1993). Anatomical differences among D. ebraccatus and D. microcephalus may contribute to reproductive isolation. Differences in male traits among these two species contribute to the production of species-typical calls (Wylcinski et al 1993). Likewise, differences in ear anatomy in females alter the 'best excitatory frequency' that each species can detect (McClelland et al 1993). Unlike species adapted to higher elevations, D. ebraccatus reduces calling rates at higher temperatures (Navas 1996a) and reduces activity at lower temperatures (Navas 1996b). Diet: Adult diet is unknown. Presumably adults eat small arthropods. Tadpoles are mainly macrophagous herbivores but will also scavenge dead tadpoles (Touchon pers. comm.). Ecology: Eggs are eaten by a wide variety invertebrate and vertebrate predators, including wasps, ants, tadpoles, turtles and snakes (Roberts 1994, Donnelly and Guyer 1994, Touchon pers. comm.). Eggs are also extremely vulnerable to dessication if they are not rained on, which negatively impacts egg survival and development and leads to altered interactions with egg and larval predators (Touchon and Warkentin 2009). Tadpoles are consumed by a wide variety of invertebrate and vertebrate predators such as fish, giant water bugs, and dragonfly nymphs (Touchon and Vonesh pers. comm.) Juveniles and adults have been observed being attacked by large spiders (Touchon pers. comm, Bernal pers. comm.). Adult D. ebbracatus have been found infected with helminth (Goldberg and Bursey 2008) and trematode (Guillen-Hernandez et al 2000) parasites. Behavior and communication: Dendropsophus ebraccatus has been described as a lek-breeding anuran (Wells 2007). Both calling perches and oviposition sites used by D. ebraccatus tend to be lower than those of a common heterospecific that also lays eggs terrestrially, Agalychnis callidryas (Donnelly and Guyer 1994). Calling by D. ebraccatus can be inhibited by calling of Dendropsophus microcephalus and Dendropsophus phlebodes (Schwartz and Wells 1983). Dendropsophus ebraccatus calling is also influenced by the calls of Scinax boulengeri due to the complete overlap in the two species' frequency ranges (Wells 2007). When multiple individuals or species are calling together at ponds, D. ebraccatus will synchronize the timing of their calls to minimize overlap (Schwartz and Wells 1984a). Dendropsophus ebraccatus also produces aggressive calls towards con- and heterospecifics (Schwartz and Wells 1985). Aggressive calls are graded in their intensity (Wells and Schwartz 1984a), and males respond more strongly to aggressive calls of increasing intensity (Wells 1989). Males lower their aggressive thresholds in response to aggressive calls, further contributing to high levels of aggressive calling observed in this species (Reichert Early Online). Calling is relatively energetically inexpensive for D. ebraccatus compared to other species with higher calling rates (Bevier 1995, 1997). Females do not necessarily prefer larger males (Morris 1991), although females do prefer calls of certain frequencies (Schwartz and Wells 1994a) as well as calls with higher pulse rates and more pulses (Wells 2007). However, large choruses and calls of heterospecifics interfere with a female's ability to assess male calls, and female preference for certain frequencies breaks down in the presence of background noise (Schwartz and Wells 1984a, Ehret & Gerhardt 1980, Gerhardt and Klump 1988a, Wollerman 1999, Wollerman and Wiley 2002a). Miyamoto and Cane (1980b) provide an excellent description of pair formation in D. ebraccatus, which has pectoral (i.e., axillary) amplexus. Silent males found near calling males will attempt to intercept females approaching calling males (Miyamoto and Cane 1980a). |
 Justin Touchon  Justin Touchon  Justin Touchon  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra  Marcos Guerra       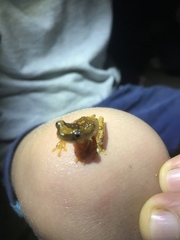                                                                    |
|
